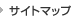




小脳を通じて脳を理解する
脳は数百億個のニューロンが複雑な神経回路網を形成して、互いに情報伝達を行うことで、その機能を実現しています。ニューロンが他のニューロンへ情報伝達を行う構造はシナプスと呼ばれ、シナプスでの情報伝達の効率の変化が、記憶や学習の細胞レベルの本体であると考えられています。そして、このシナプスの伝達効率が変化するという性質のことをシナプスの可塑性と呼びます。
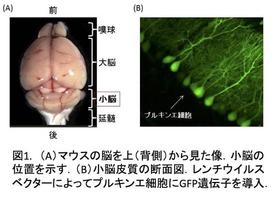
我々は記憶・学習の分子メカニズムを明らかにするために、小脳をモデルとして研究を進めてきました。小脳(図1A)は運動の巧緻性や協調性に関わる運動学習の場であり、小脳に障害が起こると運動が上手にできなくなったり、バランスが障害されたりします。運動に関係する記憶の形成には、小脳のプルキンエ細胞とそこに入力する平行線維(小脳顆粒細胞の軸索)との間のシナプスにおいて、シナプス伝達効率が長期にわたって低下することが極めて重要であることが明らかにされており、このシナプス可塑性は長期抑圧(long-term depression;LTD)と呼ばれています(図1B、図2)。
そしてLTDの分子レベルの実体は、シナプス後部表面にあるAMPA受容体の、エンドサイトーシスによる数の減少であることが分かっています。
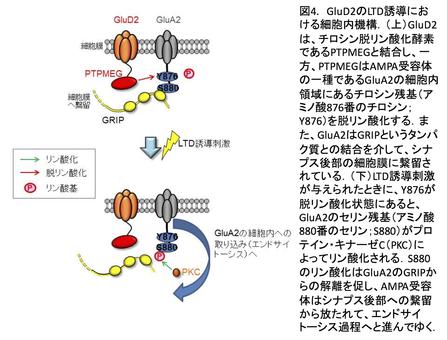
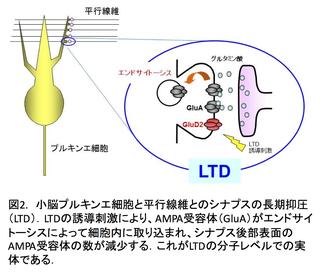
このLTDの誘導と運動学習に必須の役割を果たし、プルキンエ細胞に強く発現しているデルタ2型グルタミン酸受容体(GluD2)と平行線維から分泌されるCbln1に、我々のグループは注目しました。そしてそれらの機能的役割についての研究を進めた結果、Cbln1はシナプス前部のニューレキシン、シナプス後部のGluD2の双方に結合し、この3者の複合体形成が平行線維―プルキンエ細胞シナプスの形成・維持(図3)、そして、シナプス可塑性に必須であることを明らかにしました。さらに、GluD2からAMPA受容体のエンドサイトーシスに至るメカニズムも解明しましたが(図4)、Cbln1とGluD2の結合がどのように細胞内のシグナルをコントロールするかが、未解明なままです。そこで現在、このメカニズムを明らかにすべく、研究を進めているところです。
小脳は運動学習の場であることはよく知られていますが、ヒトでの脳の機能的イメージングや病態の研究から、小脳が認知機能にも関与していることが近年注目されています。また、ヒトにおいてGluD2が欠損すると、運動失調に加えて、言語の障害や重度の知的能力障害を呈します。マウスにおいても、従来、扁桃体や海馬が重要と考えられていた恐怖記憶に小脳が関与していることを、我々は明らかにしています。今後はさらに進んで、小脳の担う認知機能と小脳の部位や神経回路との対応付け、さらにそのメカニズムについて明らかにしてゆきたいと考えています。こうした研究を通して、古くから小脳の異常が指摘されている自閉症などの病態を分子レベル、神経回路レベルから理解できるのではないかと期待しています。